|
|
BMe Kutatói pályázat |
|

Pszichológia Doktori Iskola (Kognitív Tudomány)
BME Természettudományi Kar, Kognitív Tudományi Tanszék
Témavezető: Dr. Racsmány Mihály
A kevesebb több: miért csökkenti a figyelmi kontroll agyi hálózatának gyakorlás előtti stimulációja a sikeres tanulást?
A kutatási téma néhány soros bemutatása
Az emberi emlékezet egyes folyamatainak azonosítható agyi struktúrákhoz kötése a memóriakutatás egyik sarokköve. Az e célból végzett kutatások eszköztára egyre bővül, az idegrendszeri képalkotási technikák mellett egyre elterjedtebb a nem invazív agyi stimulációs módszerek használata, melyek még árnyaltabb képet adhatnak a memória működéséről. Kutatásunkban egy ilyen technikát (transcranial Direct Current Stimulation, azaz tDCS, koponyán keresztüli egyenáramú ingerlés) alkalmazva azt vizsgáltuk, hogy az agykéreg egy magas rendű kontrollfunkciókban érintett területének serkentő ingerlése közvetlenül a korábban elsajátított emlékek reaktivációja (felidézése, vagy az emlék tárgyával való ismételt találkozás) előtt hogyan hat ezen emlékek későbbi felidézhetőségére. Kísérleteink célja kutatócsoportunk egyik tágabb témájába illeszkedve az, hogy fényt derítsünk az emlékek stabilizálódásának módjára és ezen folyamatok agyi korrelátumaira.
A kutatóhely rövid bemutatása
A Nemzeti Agykutatási Program keretében megvalósult kutatást a BME Természettudományi Kar Kognitív Tudományi Tanszékén végeztem, dr. Racsmány Mihály Tanulás és Emlékezet Kutatócsoportjának tagjaként. Kutatócsoportunk témái közé tartozik az emlékezeti konszolidáció és előhívás, a tesztelés memóriára gyakorolt kedvező hatása, valamint a memória vizsgálata olyan kórképekben, mint a Parkinson kór és skizofrénia.
A kutatás történetének, tágabb kontextusának bemutatása
Az emberi memória empirikus vizsgálatában az utóbbi évtizedekben gyakran alkalmazott képalkotó vizsgálatok kimutatták, hogy a hosszútávú emlékezeti tárból történő felidézés egy magas szintű kognitív kontrollt igénylő folyamat [1, 2]. Ez azt jelenti, hogy viszonylag nagymértékű mentális erőfeszítésre van szükség ahhoz, hogy egy adott emléket valamilyen hívóinger (például az emlék elsajátításakor jelen levő kontextus egy része) alapján fel tudjuk idézni. Ez az erőfeszítés idegrendszeri szinten kimutatható olyan agyterületek aktivitásában, melyek érintettek a figyelmi működésekben [3, 4] – ezen területeket együttesen kontrollhálózatnak nevezzük, melynek fontos eleme a frontális lebeny. Egy emlék előhívása tehát hosszútávon (már akár percekkel az elsajátítás után) egy kontrollált keresési folyamat eredménye, mely frontális aktivitással jár. Mi történik azonban akkor, ha egy emléket ismételten „aktív” állapotba hozunk, reaktiváljuk? Több kutatás kimutatta, hogy egy emlék ismételt előhívása a frontális aktivitás folyamatos csökkenésével jár [5, 6]. Fontos kiemelni, hogy a csökkenés mértéke pozitív összefüggésben áll az emlék későbbi felidézésének sikerességével [7]. A kontrollhálózat aktivitásának csökkenéséből arra következtethetünk, hogy az előhívás a többszöri reaktiváció eredményeként egyre kevesebb kontrollt igényel, mely Racsmány és munkatársai [8] szerint azt tükrözi, hogy az előhívás folyamata egyre automatikusabbá válik – minél automatikusabb az előhívás, annál eredményesebben tudjuk felidézni az emléket hosszútávon.
Elmondhatjuk tehát, hogy az emlékek reaktivációja alatti frontális lebenyi aktivitás csökkenésének kulcsfontosságú szerepe van az emlékek megerősödésében.
A kutatás célja, a megválaszolandó kérdések
A frontális lebeny egy alterülete a dorzolaterális prefrontális kéreg (dorsolateral prefrontal cortex, DLPFC; 1. ábra), mely a kognitív kontrollhálózat egyik kiemelt fontosságú eleme [9]. Jelen kutatás azt hivatott megvizsgálni, hogy ezen terület serkentő jellegű ingerlése közvetlenül az emléknyomok reaktiválása előtt hogyan befolyásolja azok hosszútávú megtartását (felidézhetőségét). Kiindulópontunk az volt, hogy az emléknyomok ismételt reaktivációja során a kontrollhálózat (beleértve a DLPFC-t) aktivitásának fokozatos csökkenése figyelhető meg [5]. Mivel a DLPFC aktivitásának csökkenése együtt jár az emléknyomok megerősödésével, aktivitásának fenntartása várhatóan akadályozza a megerősödést elősegítő folyamatokat. A reaktiváció előtti serkentő ingerléssel tehát mintegy meggátolhatjuk a DLPFC reaktiváció alatt normálisan bekövetkező aktivitáscsökkenését, ami feltételezésünk szerint rosszabb hosszútávú emlékezeti teljesítményhez vezet.
Ugyanakkor, ha a serkentő ingerlést nem a reaktivációs periódus előtt alkalmazzuk, feltételezhetően nem lesz megfigyelhető a teljesítmény romlása. Amennyiben eredményeink alátámasztják feltételezéseinket, arra következtethetünk, hogy a DLPFC aktivitáscsökkenése az emlékek ismételt reaktivációja alatt elengedhetetlen a reaktivált emlékek megerősödéséhez.
1. ábra: A dorzolaterális prefrontális kéreg elhelyezkedése a frontális lebenyben
Módszerek
Kísérleteinkben a
DLPFC serkentő ingerlését egyenáramú ingerlés (tDCS) alkalmazásával értük el,
mely bizonyítottan alkalmas az ingerelt agyi terület aktivitásának fokozására
[10]. A módszer keretében két fejre rögzített elektróda között folyó
egyenárammal érjük el az agykéregben található idegsejtek aktivitásának
fokozását, serkentve az ingerelt agyterület aktivitását (2. ábra). A tDCS
által előidézett serkentő ingerlést a pozitív pólusú, anodális elektróda éri
el. Az anodális elektróda kísérleteinkben a jobb oldali DLPFC feletti
területen
került rögzítésre,
az ingerlés időtartama 15 perc, intenzitása 2 mA volt.
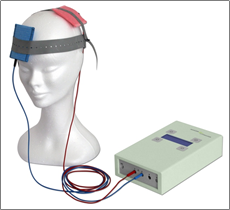
2. ábra: A tDCS gép és elektródái
A kísérletek menete
Feltevéseink vizsgálatára két kísérletet végeztünk el, mindkettő két alkalomból állt. A kísérletek számítógépes felületen zajlottak, pontos menetüket a 3. ábra szemlélteti.

Az első alkalom két részre tagolódott, a kezdeti tanulás és a reaktiváció szakaszokra, melyeket egy 30 perces késleltetés választott el.
Kezdeti tanulás
Ebben a szakaszban a kísérleti személyeknek 40 szuahéli-magyar szópárt (pl. mashua – csónak) mutattunk be egyesével a képernyőn, így az ingeranyag minden résztvevő számára egyformán újszerű volt. A tanulás 5 körön keresztül zajlott, a résztvevők feladata a szópárok memorizálása volt.
30 perc késleltetés
A késleltetés elején felhelyeztük az elektródákat, ezt követően a hátralevő időben a kísérleti személyek aritmetikai feladatokat oldottak meg, melynek célja a megtanult szavak magukban történő felmondásának megakadályozása volt.
Reaktiváció
A késleltetés után a reaktivációs szakasz következett, melyben a résztvevők a kezdeti tanulás során megtanult szópárok felét ismételt teszteléssel, másik felüket ismételt tanulással gyakorolták, 5 körön keresztül. A teszteléssel történő gyakorlás keretében a szuahéli szó láttán kellett felidézniük a magyar szót, és begépelniük azt. Újratanulás esetén a teljes szópár bemutatásra került, csakúgy, mint a kezdeti tanulási szakaszban. Ilyen módon tehát az ismételt tesztelés és az ismételt tanulás jelentették a reaktiváció kétféle módját.
A kísérletek második alkalmára egy héttel később került sor, ekkor a végső teszt során résztevők feladata az összes szópár magyar felének egyszeri felidézése volt, a szuahéli szó láttán.
Két kísérletünk elrendezésében az egyetlen különbség az volt, hogy az 1. kísérletben a tDCS általi serkentő ingerlésre közvetlenül a reaktivációs fázis előtt, a 30 perces késleltetés utolsó 15 percében került sor, a 2. kísérletben pedig az ingerlés közvetlenül a reaktivációs fázis utáni 15 percben zajlott. A résztvevőket mindkét kísérletben két csoportra osztottuk. A kísérleti csoportban a résztvevők valódi, 15 percen át tartó anodális ingerlésben részesültek (anodális csoport), míg a kontroll csoportban csupán néhány másodpercig tartó, úgynevezett sham ingerlést kaptak (sham csoport), mely nem indukál a hosszabb ingerléssel összevethető változást az idegsejtek működésében.
A reaktivációs szakaszban és a végső teszt alatt két változót mértünk, ezek mentén hasonlítottuk össze a két ingerlési csoportot: a helyesen felidézett szavak száma (felidézési teljesítmény), és a helyesen felidézett szavak előhívásának gyorsasága (reakcióidő).
Eddigi eredmények
1. kísérletünk legfontosabb eredménye, hogy a hosszútávú felidézési teljesítmény tekintetében szignifikáns különbség volt kimutatható az anodális és sham csoportok között. Ez azt jelenti, hogy a végső teszt során az anodális csoport kevesebb szópárt tudott felidézni, mint a sham csoport, több mint 10%-os különbség volt megfigyelhető (4. ábra). A reakcióidő tekintetében nem volt különbség a két csoport között.
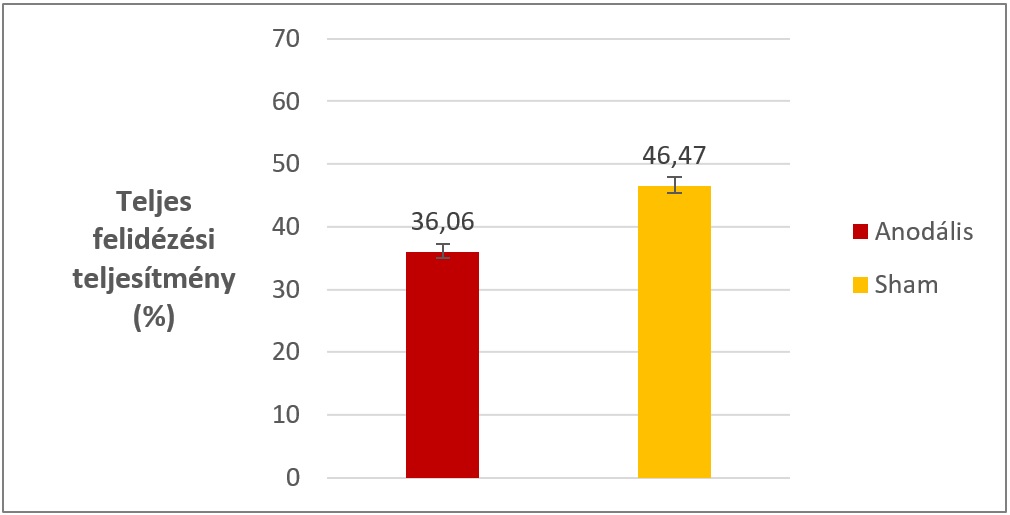
4.
ábra: A két ingerlési csoport teljes felidézési teljesítménye az 1. kísérlet
végső tesztjén. A különbség szignifikáns. Hibasávok: standard hiba
Fontos, hogy az anodális ingerlés hatása mind az ismételt teszteléssel, mind az ismételt tanulással reaktivált emlék esetén kimutatható volt, a hatás tehát független a reaktiváció típusától. Szintén fontos kiemelni, hogy a két ingerlési csoport között rövidtávon, azaz a reaktivációs periódus alatt sem a felidézési teljesítmény, sem a reakcióidő tekintetében nem volt különbség: a teszteléssel történő reaktiváció során mindkét csoport azonos mennyiségű szópárt idézett fel, azonos sebességgel, ez igaz volt mindkét reaktiváció típus esetén.
1. kísérletünk eredményei arra engednek következtetni, hogy a DLPFC-n a reaktiváció előtt alkalmazott serkentő ingerlés hosszútávú negatív hatást gyakorol a reaktivált emlékek felidézhetőségére. Az ugyanakkor nem eldönthető, hogy az ingerlés valóban a reaktiváción fejtette-e ki hatását, hiszen az ingerlés időben a kezdeti tanuláshoz is közel állt, így nem zárhatjuk ki, hogy a tanulás utáni emlékezeti konszolidációs folyamatokra volt hatással. Ennek megválaszolására végeztük el 2. kísérletünket, melyben az ingerlés a reaktivációs periódust követően zajlott, ahol feltételezhetően szintén konszolidációs folyamatok zajlanak.
2. kísérletünk legfőbb eredménye, hogy hosszútávon sem a felidézési teljesítmény, sem a reakcióidő tekintetében nem volt különbség a két ingerlési csoport között, ami azt jelenti, hogy a végső teszt során mindkét csoport közel egyenlő számú szópárt tudott felidézni, közel azonos sebességgel (X. ábra). Elvárásainknak megfelelően rövidtávon, a reaktivációs szakaszban sem volt különbség a két ingerlési csoport között a felidézési teljesítményben, vagy a reakcióidőkben, hiszen az ingerlés a reaktiváció után történt.
2. kísérletünk eredményeiből arra következtethetünk, hogy az ingerlés szelektíven a reaktivációs perióduson fejtette ki hatását (ahogyan az 1. kísérletben láttuk), hiszen az utána adott ingerlés nem volt hatással az emlékek felidézhetőségére.
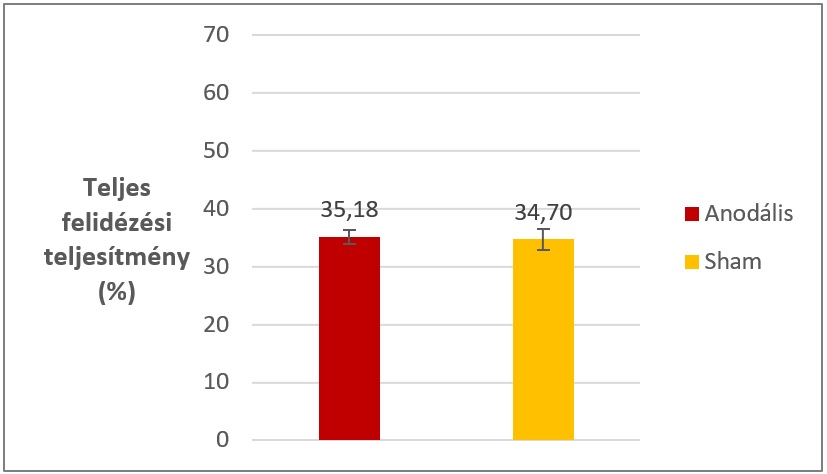
5. ábra: A két ingerlési csoport teljes felidézési teljesítménye a 2. kísérlet végső tesztjén. A különbség nem szignifikáns. Hibasávok: standard hiba
Mindkét kísérletünkben megfigyelhető, hogy az ismételt tesztelés az újratanuláshoz képest előnyösebb gyakorlási stratégia, mely jobb hosszútávú megtartáshoz vezet: az újratesztelt szópárokat hosszútávon szignifikánsan nagyobb sikerrel és gyorsabban tudták felidézni a résztvevők, mint az újratanultakat; ez a jellemző volt mindkét kísérletünkben (x. ábra). A tesztelés előnyös hatása az újratanulással szemben a szakirodalomban a tesztelési hatás néven ismert [11], új eredményeink mellet tehát egy már jól megalapozott hatást is replikáltunk.
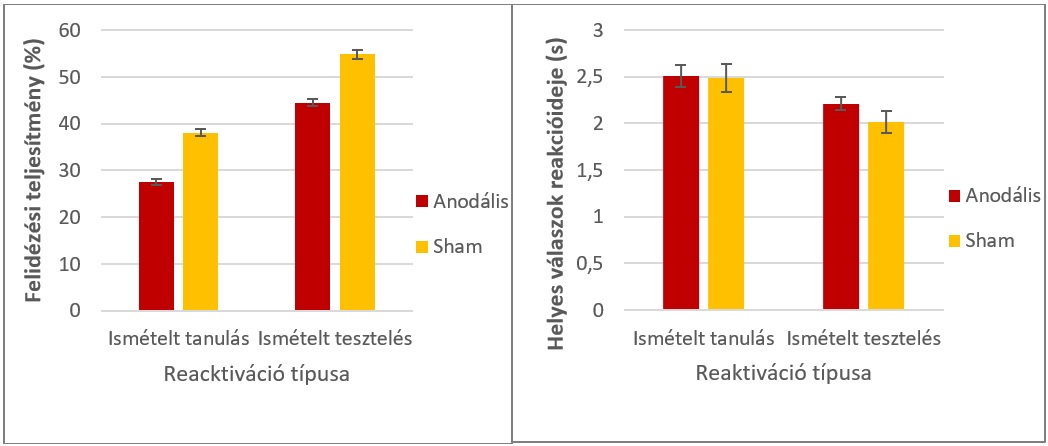
6. ábra: A felidézési teljesítmény (bal) és a reakcióidők (jobb) az 1. kísérlet végső tesztje során. A tesztelés sikeresebb és gyorsabb hosszútávú felidézéshez vezet. A 2. kísérletben hasonló mintázatot figyeltünk meg. Hibasávok: standard hiba
Összességében elmondhatjuk, hogy a DLPFC serkentő jellegű ingerlése az emlékek reaktivációja előtt nincs hatással a rövidtávú memória teljesítményére, de negatívan befolyásolta a reaktivált emlékek stabilizálódását (a reaktiváció típusától függetlenül), ami a hosszútávú felidézési teljesítmény romlásában mutatkozik meg.
Várható impakt, további kutatás
Saját publikációk, hivatkozások
Kapcsolódó saját publikációk listája
Poszterek és előadások hazai és nemzetközi konferenciákon
Marián M., Szőllősi Á., Zimmer M., Racsmány M. (2016). Anodal transcranial stimulation of right prefrontal cortex deteriorates long term memory consolidation. Poszter. International Conference on Memory. Budapest, Magyarország, 2016. július 17-22.
Szőllősi Á., Marián M., Zimmer M., Racsmány M. (2017). The detrimental impact of offline transcranial direct current stimulation of the right prefrontal cortex on long-term verbal associative memory performance. Poszter. International Brain Stimulation Conference. Barcelona, Spanyolország, 2017. március 5-8.
Marián M., Szőllősi Á., Zimmer M., Racsmány M. (2017). A jobb oldali prefrontális kéreg egyenáram-ingerlése csökkenti a reaktivált emlékek konszolidációját. Előadás. A Magyar Pszichológiai Társaság XXVI. Országos Nagygyűlése. Szeged, Magyarország, 2017. június 1-3.
Marián M., Szőllősi Á., Zimmer M., Racsmány M. (2017). Direct current stimulation of right prefrontal cortex decreases consolidation of reactivated memories. Előadás. A BME Kognitív Tudományi Tanszék éves PhD Konferenciája. Budapest, Magyarország, 2017. június 16.
Hivatkozások listája
2. Nyberg, L., Cabeza, R., & Tulving, E. (1996). PET studies of encoding and retrieval: The HERA model. Psychonomic Bulletin & Review, 3, 135-148.
3. Cabeza, R., Dolcos, F., Graham, R., & Nyberg, L. (2002). Similarities and differences in the neural correlates of episodic memory retrieval and working memory. NeuroImage, 16, 317–30.
4. Ranganath, C., Johnson, M. K. & D’Esposito, M. (2003). Prefrontal activity associated with working memory and episodic long-term memory. Neuropsychologia, 41, 378–389.
5. Kuhl, B. A., Dudukovic N. M., Kahn, I. & Wagner, A. D. (2007). Decreased demands on cognitive control reveal the neural processing benefits of forgetting. National Neuroscience, 10, 908–914.
6. Keresztes, A., Kaiser, D., Kovács, G. & Racsmány, M. (2014). Testing promotes long-term learning via stabilizing activation patterns in a large network of brain areas. Cerebral Cortex, 24, 3025–3035.
7. Eriksson, J., Kalpouzos, G. & Nyberg, L. (2011). Rewiring the brain with repeated retrieval: A parametric fMRI study of the testing effect. Neuroscience letters, 505, 36-40.
8. Racsmány, M., Szőllősi, Á. & Bencze, D. (2017). Retrieval Practice Makes Procedure From Remembering: An Automatization Account of the Testing Effect. Journal of Experimental Psychology: Learning, Memory and Cognition. Advance online publication.
9. Blumenfeld, R. S. & Ranganath, C. (2006). Dorsolateral Prefrontal Cortex Promotes Long-Term Memory Formation through Its Role in Working Memory Organization. The Journal of Neuroscience, 3, 916 –925.
10. Au, J., Katz, B., Buschkuehl, M., Bunarjo, K., Senger, T., Zabel, C., Jaeggi, S. M. & Jonides, J. (2016). Enhancing Working Memory Training with Transcranial Direct Current Stimulation. Journal of Cognitive Neuroscience, 9, 1419–1432.
11. Roediger, H.L., & Karpicke, J.D. (2006). Test enhanced learning: Taking memory tests improves long-term retention. Psychological Science, 17, 249–255.