|
|
BMe Research Grant |
|

Doctoral School of Psychology (Cognitive Science)
BME TTK, Department of Cognitive Science
Supervisor: Dr. Racsmány Mihály
Less is More: Why Does Stimulation of the Attentional Neural Network before Practice Decrease Success of Learning?
Introducing the research area
Identifying brain areas involved in specific memory processes is a cornerstone of memory research. The toolbox of research is expanding, and besides neuroimaging techniques, use of non-invasive stimulation methods is becoming frequent. These can provide us with important insight into memory functioning. Using such a method (transcranial Direct Current Stimulation, tDCS) we investigated whether excitatory stimulation of a brain area involved in high order control functions immediately before reactivation (recall or re-presentation) of memories can affect the recall success of the memories in the long term. The aim of this study, fitting with the broader goal of one of the topics of our research team is to shed light on the process of memory stabilization and its neural correlates.
Brief introduction of the research place
The study, supported by the Hungarian Brain Research Program was conducted at the Department of Cognitive Science, at the Faculty of Natural Sciences. I conducted the study as a member of the Learning and Memory Research Group, led by Mihály Racsmány, PhD. The topics of our research group include memory encoding and consolidation, the beneficial effects of testing, and investigation of memory in Parkinson’s disease and schizophrenia.
History and context of research
Empirical investigations, predominantly neuroimaging studies of memory concepts have shown that retrieving a long-term memory requires high order cognitive control [1, 2]. This means mental effort is needed to recall a memory upon a cue (e.g. part of the context present at encoding). This effort is mirrored in neural activity in areas involved in attentional processes [3, 4] – collectively known as the control network, and an important part of this network is the frontal lobe. Thus, retrieval of a memory in the long term (even minutes after encoding) is the result of a controlled search process, reflected in frontal activity. But what happens if we repeatedly return a memory into an “active” state, reactivate it? Multiple recent studies have shown that repeated retrieval of a memory leads to a gradually decreasing frontal activity [5, 6]. It is important to emphasize that this decrease is positively correlated with later recall success of a given memory [7]. Decreasing activity of the control network can lead us to the conclusion that retrieval, as a result of repeated reactivation requires less and less control, which according to Racsmány and colleagues [8] is paralleled by an automatization process – the more automatic retrieval is, the more successful long-term retention will be.
We can conclude that decreasing activity in frontal areas during reactivation is crucial for the stabilization of memories.
The research goal, open questions
The dorsolateral prefrontal cortex (DLPFC, Figure 1), as part of the frontal lobe, is an important component of the control network [9]. Present study aimed to investigate how excitatory stimulation of this area immediately before reactivation of memories affects long term retention of these memories. Our starting point was that repeated reactivation of memories is accompanied by decreasing control network activity (including DLPFC) [5]. Since decreasing activity in the DLPFC is correlated with stabilization of memories, maintaining its activity will presumably hinder processes normally beneficial to stabilization. Excitatory stimulation before reactivation can thus inhibit the normal decrease of DLPFC activity during reactivation which will result in a worsened long-term recall performance. At the same time, if excitatory stimulation is carried out at a different point (i.e. not before reactivation), we expect no such worsening. In case our results are in line with our assumptions, we can conclude that activity decrease in the DLPFC during reactivation is crucial for stabilization of memories.
Figure
1. Location of the
dorsolateral
prefrontal cortex within the frontal lobe
Methods
Characteristics of tDSC
Excitatory stimulation of the DLPFC was achieved by direct current stimulation (tDCS), which method is proven to effectively modify activity in the stimulated areas [10]. Direct current flows between electrodes fixed on the scalp, modifying the activity of neurons, exciting or inhibiting a neural region (Figure 2). Excitatory stimulation is achieved by the positive, anodal electrode. In our experiments, the anodal electrode was fixed over the right DLPFC, the duration of stimulation was 15 minutes, and intensity was 2 mA.
Figure
2. tDCS
equipment
Experimental procedure
To
investigate our assumptions, we conducted two experiments, both consisting of
two sessions. Experiments were carried out on a computer platform; exact
procedures can be seen in Figure 3.
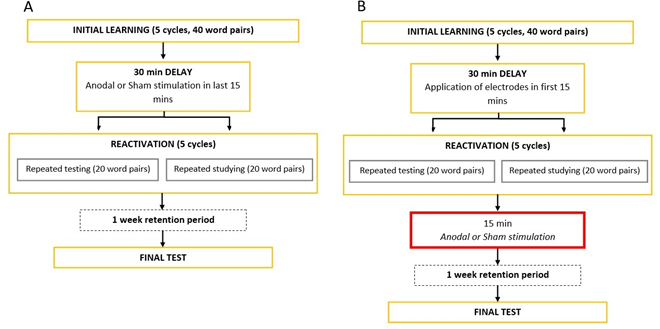
Figure 3. Experimental procedures. A: Experiment
1. B: Experiment 2.
The first session consisted of two phases, initial learning and reactivation, which were separated by a 30-minute delay.
Initial learning
In this phase, subjects learned 40 Swahili-Hungarian word pairs (e.g. mashua – csónak), so that the material was equally novel for all the participants. Subjects learned all 40 word pairs in 5 cycles.
30-minute delay
During the delay, tDCS electrodes were applied and subjects solved arithmetic tasks to prevent them from rehearsing the studied material.
Reactivation
Delay was followed by the reactivation phase, where subjects practiced half the word pairs by repeated testing and the other half by repeated studying in 5 cycles. During practice by repeated testing they had to recall the Hungarian part of the pair upon the Swahili cue, and type it in. During repeated study, word pairs were repeatedly presented as a whole. Thus, repeated testing and repeated studying were the two types of reactivation.
The second session took place exactly one week later, when subjects were required to recall all word pairs (Hungarian word upon the Swahili cue) in the final test phase.
The only difference between the two experiments was that in Experiment 1, excitatory stimulation of the DLPFC took place immediately before the reactivation phase, in the last 15 minutes of the 30-minute delay, whereas in Experiment 2, stimulation was executed immediately after the reactivation phase. Subjects in both experiments were divided into two groups. The experimental group received actual 15-minute anodal stimulation (anodal group), while the control group only received a stimulation of a few seconds (sham group), which does not induce effects comparable to a longer stimulation duration.
In our analysis, we compared the two stimulation groups along two variables measured in the reactivation and final test phases: number of correctly recalled words (recall success) and reaction time of correctly recalled items.
Results
The key finding of Experiment 1 is that there was a significant difference in recall success between anodal and sham groups at the final test; the anodal group performed about 10% worse (Figure 4.). There was no such difference in reaction times. Importantly, the detrimental effect of anodal stimulation was observable for both reactivation types.
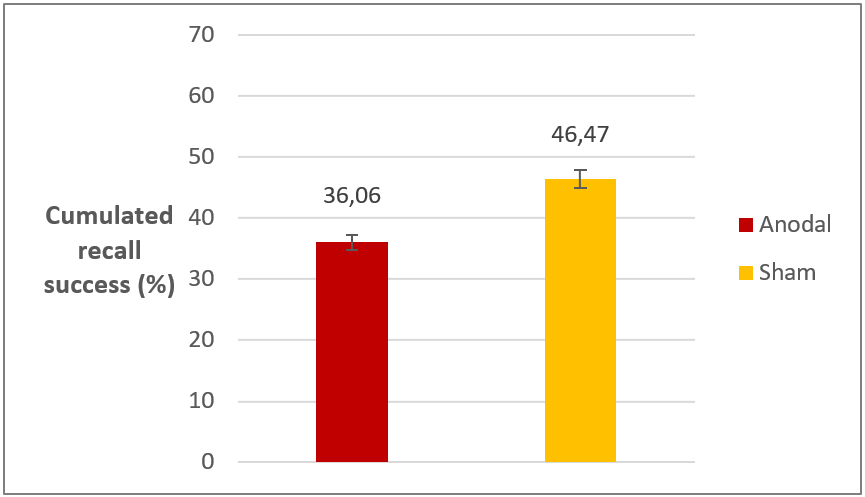
Figure 4. Recall performance on
the final test in Experiment 1 in the two stimulation groups. Difference
is significant. Error bars represent standard error.
It is also important to emphasize that in the short term (in the reactivation phase) there was no difference between the two stimulation groups either in recall success or reaction times.
Based on these results we can conclude that excitatory stimulation of the DLPFC immediately before memory reactivation had a negative effect on the recall of these memories. However, we cannot determine whether stimulation truly had an effect on reactivation or on consolidation processes taking place after the initial learning during the delay. For this reason, we conducted a second experiment, where stimulation was carried out after reactivation, where consolidation processes are also thought to occur.
The main results of Experiment 2 are that at the final test there was no difference between the stimulation groups in either recall success or reaction times (figure 5). In line with our expectations, stimulation groups did not differ in the short term either. Based on these results we can conclude that anodal stimulation selectively affected the reactivation period (as seen in the Experiment 1), since stimulation after reactivation had no effect on memory performance.
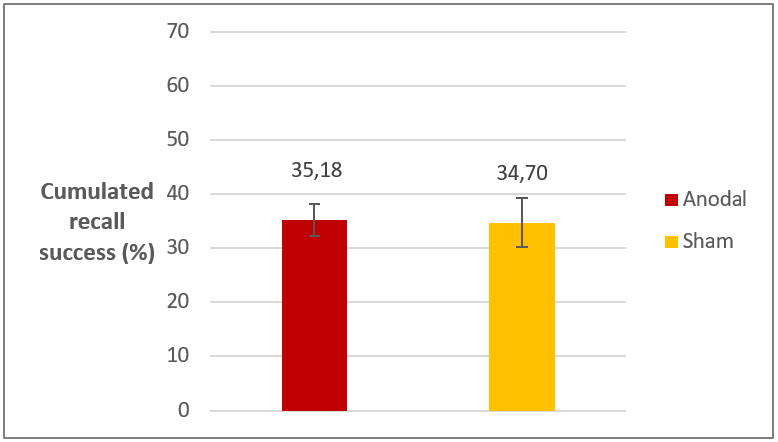
Figure 5. Recall performance on the final test in Experiment 2 in the two stimulation groups. Difference is not significant. Error bars represent standard error.
Besides our novel findings, in both experiments we replicated the well-known phenomenon of the testing effect [11], according to which repeated testing of a material is a more efficient practice strategy than repeated studying: word pairs practiced by testing showed a significantly better retention and faster reaction times than repeatedly studied ones (Figure 6).
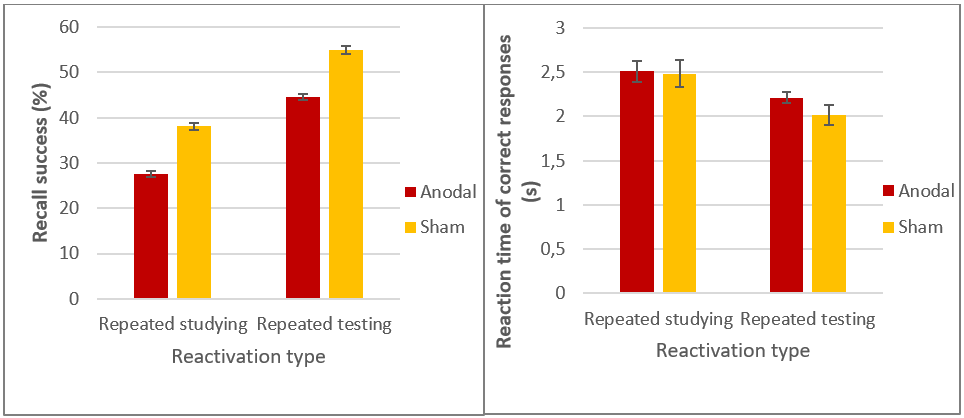
Figure 6. Recall performance (left) and reaction times (right) in the final test of Experiment 1. Practice by testing leads to more successful and faster memory performance. In Experiment 2 we found a similar pattern. Error bars represent standard error.
Overall, we can conclude that excitatory stimulation of the DLPFC immediately before memory reactivation has no effect on short term memory performance, but negatively affects stabilization of the memories (irrespective of the type of reactivation), which reflects in a worsening of long-term retention.
Expected impact and further research
Our results point to the conclusion that exciting the DLPFC (one of the main areas of cognitive control) during memory reactivation has seemingly paradoxical negative effects on the retention of memories. From this we can infer that in line with previous results [6] the normally occurring decrease in activity of the DLPFC has an important role in the stabilization of memories. One of the advantages of this study is that we investigated a long-term memory process, which, to our knowledge has not been done with tDCS before. Present study and other ongoing and planned studies in our laboratory help us understand what mechanisms are necessary for memory stabilization, that is to understand how our experiences become memories that we can retrieve in the long term.
Own publications, references
Related publications
Posters and presentations at national and international conferences
Marián M., Szőllősi Á., Zimmer M., Racsmány M. (2016). Anodal transcranial stimulation of right prefrontal cortex deteriorates long term memory consolidation. Poster. International Conference on Memory. Budapest, Hungary, 17-22 June, 2016.
Szőllősi Á., Marián M., Zimmer M., Racsmány M. (2017). The detrimental impact of offline transcranial direct current stimulation of the right prefrontal cortex on long-term verbal associative memory performance. Poster. International Brain Stimulation Conference. Barcelona, Spain, 5-8 March, 2017
Marián M., Szőllősi Á., Zimmer M., Racsmány M. (2017). A jobb oldali prefrontális kéreg egyenáram-ingerlése csökkenti a reaktivált emlékek konszolidációját. Presentation. A Magyar Pszichológiai Társaság XXVI. Országos Nagygyűlése. Szeged, Hungary, 1-3 June, 2017
Marián M., Szőllősi Á., Zimmer M., Racsmány M. (2017). Direct current stimulation of right prefrontal cortex decreases consolidation of reactivated memories. Presentation. Yearly PhD Conference of the Department Of Cognitive Science of BME. Budapest, Hungary, 16 June, 2017
References
1. Badre, D. & Wagner., A. D. (2007). Left ventrolateral prefrontal cortex and the cognitive control of memory. Neuropsychologia, 45, 2883–2901.
2. Nyberg, L., Cabeza, R., & Tulving, E. (1996). PET studies of encoding and retrieval: The HERA model. Psychonomic Bulletin & Review, 3, 135-148.
3. Cabeza, R., Dolcos, F., Graham, R., & Nyberg, L. (2002). Similarities and differences in the neural correlates of episodic memory retrieval and working memory. NeuroImage, 16, 317–30.
4. Ranganath, C., Johnson, M. K. & D’Esposito, M. (2003). Prefrontal activity associated with working memory and episodic long-term memory. Neuropsychologia, 41, 378–389.
5. Kuhl, B. A., Dudukovic N. M., Kahn, I. & Wagner, A. D. (2007). Decreased demands on cognitive control reveal the neural processing benefits of forgetting. National Neuroscience, 10, 908–914.
6. Keresztes, A., Kaiser, D., Kovács, G. & Racsmány, M. (2014). Testing promotes long-term learning via stabilizing activation patterns in a large network of brain areas. Cerebral Cortex, 24, 3025–3035.
7. Eriksson, J., Kalpouzos, G. & Nyberg, L. (2011). Rewiring the brain with repeated retrieval: A parametric fMRI study of the testing effect. Neuroscience letters, 505, 36-40.
8. Racsmány, M., Szőllősi, Á. & Bencze, D. (2017). Retrieval Practice Makes Procedure From Remembering: An Automatization Account of the Testing Effect. Journal of Experimental Psychology: Learning, Memory and Cognition. Advance online publication.
9. Blumenfeld, R. S. & Ranganath, C. (2006). Dorsolateral Prefrontal Cortex Promotes Long-Term Memory Formation through Its Role in Working Memory Organization. The Journal of Neuroscience, 3, 916 –925.
10. Au, J., Katz, B., Buschkuehl, M., Bunarjo, K., Senger, T., Zabel, C., Jaeggi, S. M. & Jonides, J. (2016). Enhancing Working Memory Training with Transcranial Direct Current Stimulation. Journal of Cognitive Neuroscience, 9, 1419–1432.
11. Roediger, H.L., & Karpicke, J.D. (2006). Test enhanced learning: Taking memory tests improves long-term retention. Psychological Science, 17, 249–255.